 Catalogue PIGMA
Catalogue PIGMA
CSV
Type of resources
Available actions
Topics
Keywords
Contact for the resource
Provided by
Years
Formats
Representation types
Update frequencies
status
Scale
Resolution
-
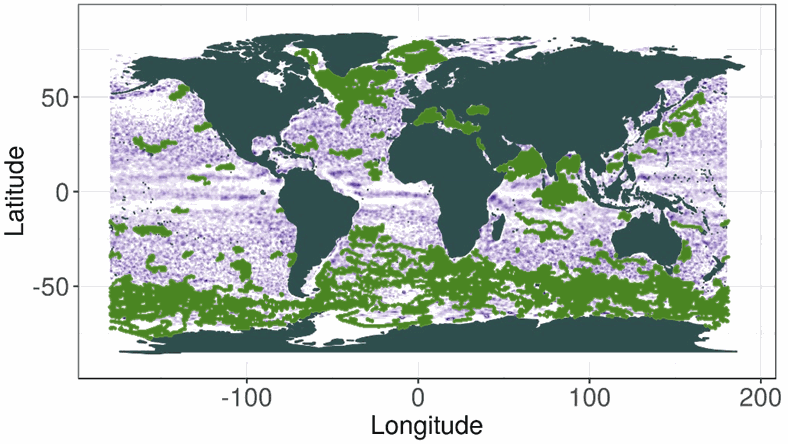
This dataset contains bio-optical measurements and environmental parameters associated with Deep Chlorophyll Maxima acquired by BGC-Argo profiling floats and matched with mesoscale eddies location from the Ocean Eddy Detection and Tracking Algorithms (TOEeddies) Atlas. For each BGC-Argo profile the data files includes the World Meteorological Organistion (WMO) and profile numbers, geographical position (LON and LAT), the date of the profile in Julian Day (JULD); the qualification of the vertical profile (CARAC_BIO) as Deep Biomass Maximum (DBM), Deep photoAcclimation Maximum (DAM), or presenting no DCM (NO); at the depth of the maximum (DCM_DEPTH), the chlorophyll a concentration (CHLA_DCM, mg chla m-3 ) and the backscattering coefficient (BBP_DCM, m-1); the Mixed Layer Depth (MLD, m), the nitracline depth (NCLINE, m), the mean daily Available PAR in the Mixed Layer Depth (MIPAR_MLD, E m -1 d -1), the daily Available PAR at the nitracline depth (IPAR_NCLINE, E m-2 d-1); the location of the profile (CARAC_EDDY) as being inside the core/periphery (IN/_EN) of a cyclonic/anticyclonic eddy (DEP_/P_), or outside eddy influence (OUT); the processing level (MODE) of the ADT maps used for the TOEddies detection, either Near Real Time (-1), or Delayed Mode (1). The qualification and processing of the BGC-Argo profiles, as well as the DCM detection (DAM/DBM/NO) and the estimation of the environmental parameters, were applied as described from Cornec, M., Claustre, H., Mignot, A., Guidi, L., Lacour, L., Poteau, A., D'Ortenzio, F., Gentili, B., Schmechtig, C. (2021). Deep chlorophyll maxima in the global ocean: Occurrences, drivers and characteristics. Global Biogeochem Cycles 35. https://doi.org/10.1029/2020GB006759 Data relative to mesoscale eddies were produced by processing daily 0.25°x0.25° AVISO Absolute Dynamical Topography (maps produced by Ssalto/Duacs and distributed by Copernicus-Marine Environment Services) with the TOEddies algorithm (Laxenaire, R., Speich, S., Blanke, B., Chaigneau, A., Pegliasco, C., & Stegner, A. (2018). Anticyclonic Eddies Connecting the Western Boundaries of Indian and Atlantic Oceans. Journal of Geophysical Research: Oceans, 123(11), 7651–7677. https://doi.org/10.1029/2018JC014270)
-
Diet and stable isotopes of the European eel (Anguilla anguilla L.) in six northern France estuaries
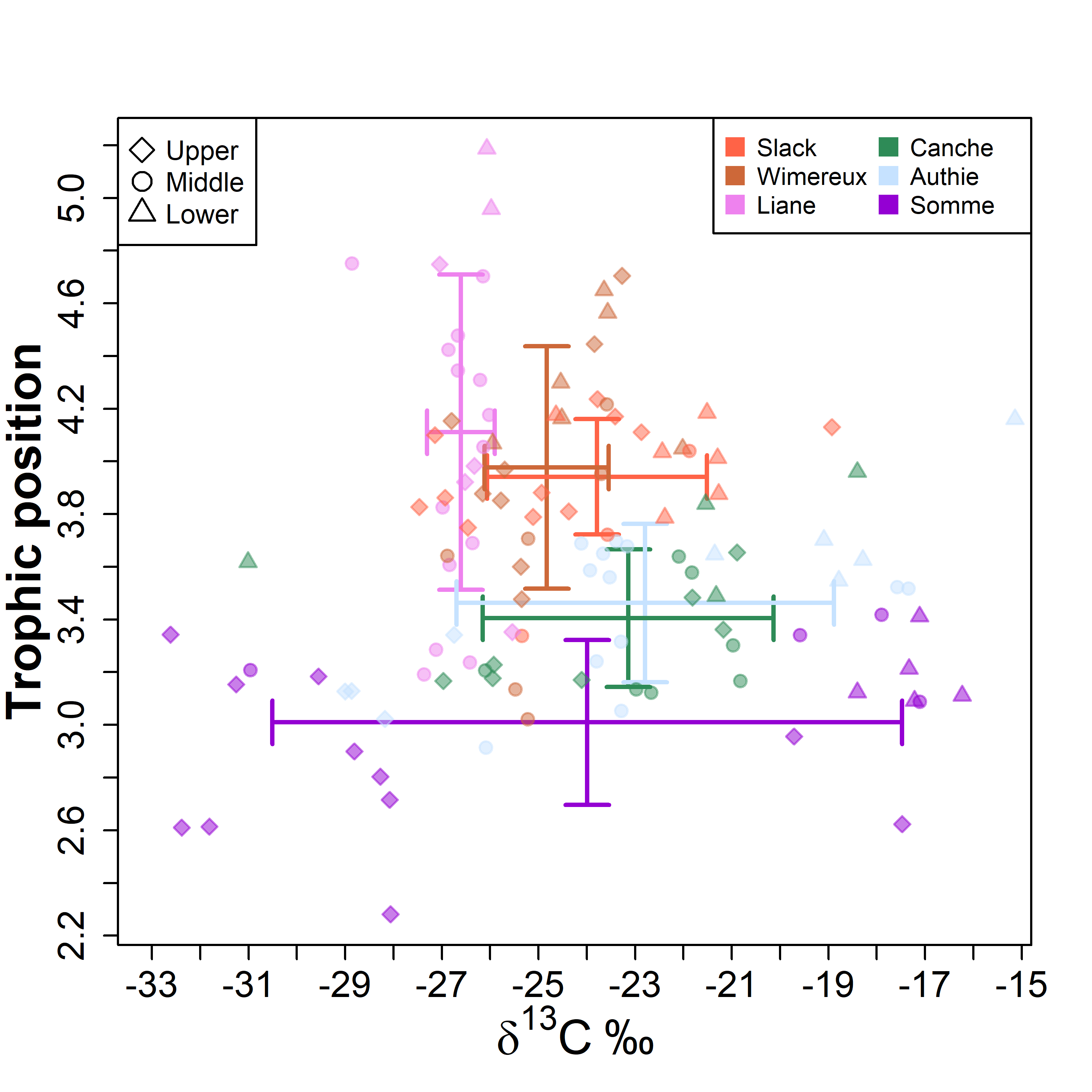
The diet and stable isotopic (i.e. δ15N and δ13C values) compositions of eels have been studied during each season of 2019 with a fyke net in six estuaries located along the French coast of the eastern English Channel (Slack, Wimereux, Liane, Canche, Authie and Somme estuaries) (10.1371/journal.pone.0270348).
-
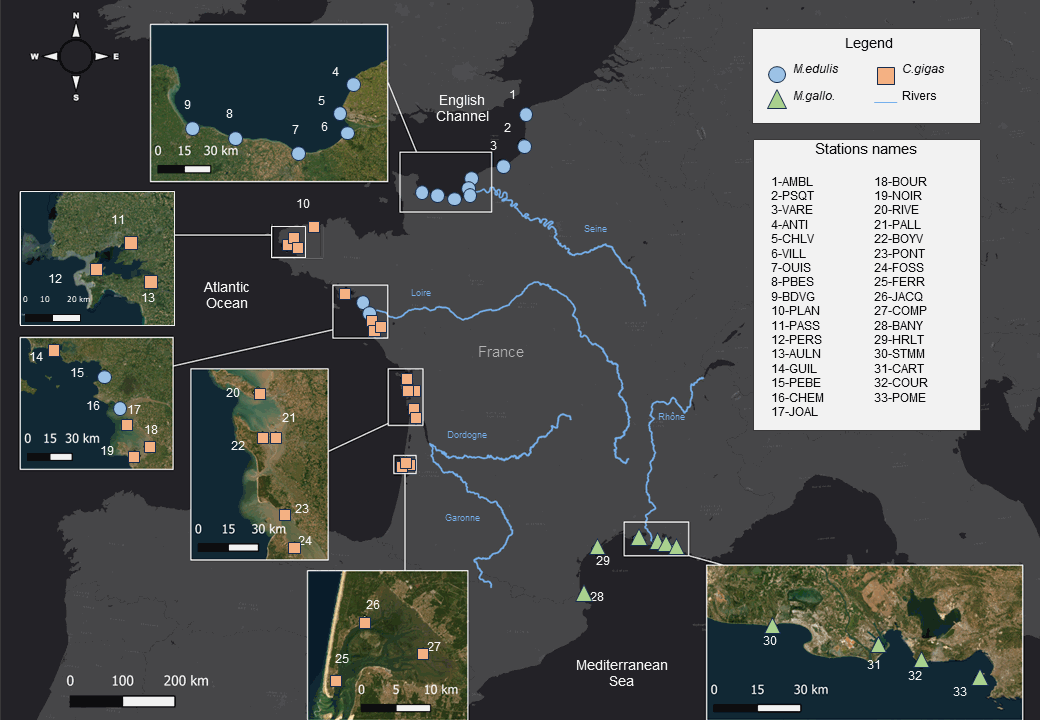
Bivalves carbon and nitrogen elemental and isotopic ratios (δ13C, δ15N, C and N%, C:N) times series (1981-2021) from 33 sites in France. Bivalve species are the Pacific oyster Crassostrea gigas, and the mussels Mytilus edulis and Mytilus galloprovincialis. This extensive dataset offers a comprehensive view spanning multiple decades and ecosystems, allowing to track how coastal ecosystems and marine species record changing climate, physical-chemical environments and organic matter cycles. This dataset may also be used to study bivalve physiology. Additionally, these data are crucial for establishing isotope baselines for studying food webs. Ultimately, this data set provide valuable information for more effective ecosystem conservation and management strategies in our rapidly changing world.
-
Aires géographiques des appellations d'origine contrôlées (AOC)/protégées (AOP). Le fichier liste pour chaque commune, identifiée par son département, son nom et son code INSEE, les aires géographiques des appellations AOC/AOP qui se situent sur la commune
-

In October 2019 we chose 15 sites from the 2019 EVHOE survey for environmental DNA (eDNA) sampling. The French international EVHOE bottom trawl survey is carried out annually during autumn in the BoB to monitor demersal fish resources. At each site, we sampled seawater using Niskin bottles deployed with a circular rosette. There were nine bottles on the rosette, each of them able to hold ∼5 l of water. At each site, we first cleaned the circular rosette and bottles with freshwater, then lowered the rosette (with bottles open) to 5 m above the sea bottom, and finally closed the bottles remotely from the boat. The 45 l of sampled water was transferred to four disposable and sterilized plastic bags of 11.25 l each to perform the filtration on-board in a laboratory dedicated to the processing of eDNA samples. To speed up the filtration process, we used two identical filtration devices, each composed of an Athena® peristaltic pump (Proactive Environmental Products LLC, Bradenton, Florida, USA; nominal flow of 1.0 l min–1 ), a VigiDNA 0.20 μm filtration capsule (SPYGEN, le Bourget du Lac, France), and disposable sterile tubing. Each filtration device filtered the water contained in two plastic bags (22.5 l), which represent two replicates per sampling site. We followed a rigorous protocol to avoid contamination during fieldwork, using disposable gloves and single-use filtration equipment and plastic bags to process each water sample. At the end of each filtration, we emptied the water inside the capsule that we replaced by 80 ml of CL1 conservation buffer and stored the samples at room temperature following the specifications of the manufacturer (SPYGEN, Le Bourget du Lac, France). We processed the eDNA capsules at SPYGEN, following the protocol proposed by Polanco-Fernández et al., (2020). Half of the extracted DNA was processed by Sinsoma using newly developped ddPCR assays for European seabass (Dicentrachus labrax), European hake (Merluccius merluccius) and blackspot seabream (Pagellus bogaraveo). The other half of the extracted DNA was analysed using metabarcoding with teleo primer. The raw metabarcoding data set is available at https://www.doi.org/10.16904/envidat.442 Bottom trawling using a GOV trawl was carried out before or after water sampling. The catch was sorted by species and catches in numbers and weight were recorded. No blackspot seabream individuals were caught. Data content: * ddPCR/: contains the ddPCR counts and DNA concentrations for each sample and species. * SampleInfo/: contains the filter volume for each eDNA sample. * StationInfo/: contains metadata related to the data collected in the field for each filter. * Metabarcoding/: contains metabarcoding results for teleoprimer. * Trawldata/: contains catch data in numbers and weight (kg).
-

This REPHY dataset includes long-term time series on marine phytoplankton and physico-chemical measures, since 1987, along the whole French metropolitan coast. Some of these data are shared with those of the following regional networks: SRN (Hauts-de-France), RHLN (Normandy), ARCHYD (Arcachon), RSLHYD (Mediterranean lagoons). REPHY dataset from overseas departments (Martinique, Guadeloupe and French Guiana in West Atlantic waters; Reunion Island and Mayotte in Indian Ocean) will be available later. Phytoplankton data essentially cover microscopic taxonomic identifications and counts, but also pigments measures and flux cytometry measures in few regions. Physico-chemical measures include temperature, salinity, turbidity, dissolved oxygen, nutrients and chlorophylle a. The whole dataset is available, but is also divided into regions: North Sea + Channel + Atlantic, Mediterranean. For each of these two regional datasets, one includes only phytoplankton counts (PHYTO), the other (HYDRO) includes physico-chemical measures, pigments and flux cytometry measures.
-

This folder contains two examples of PAGURE datasets, corresponding to three surveys: -CGFS conducted in 2018 in the English Channel (Northeast Atlantic) -EPIBENGOL conducted in 2019 in the Gulf of Lion (Western Mediterranean) -EVHOE conducted in 2020 in the Bay of Biscay and Celtic Shelf (Northeast Atlantic) Files include metadata for the sampling stations, annotation files. A readme tex file contains the links to the voyage metadata This folder is aimed at providing an example of documented underwater imagery dataset. These data are part of the data exchange conducted in the QuatreA collaboration between the French Research Institute for the Exploitation of the Sea (Ifremer), the Commonwealth Scientific and Industrial Research Organisation (CSIRO), and the University of Tasmania (UTAS).
-

As part of the marine water quality monitoring of the “Pertuis” and the “baie de l’Aiguillon” (France), commissioned by the OFB and carried out by setec énergie environnement, three monitoring stations were installed. Two of them were set up at the mouths of the Charente and Seudre rivers on February 6 and 27, 2019, respectively, while a third was deployed in the Bay of Aiguillon on March 24, 2021. The dataset presented here concerns the station installed in the Bay of Aiguillon. Measurements are organized into .csv files, with one file per year. Data is collected using a WiMO multiparameter probe, which records the following parameters: • Temperature (-2 to 35 °C) • Conductivity (0 to 100 mS/cm) • Pressure (0 to 30 m) • Turbidity (0 to 4000 NTU) • Dissolved Oxygen (0 to 23 mg/L & 0 to 250 %) • Fluorescence (0 to 500 ppb)
-
210Pb, 226Ra and 137Cs were measured by non-destructive gamma spectrometry on marine sediment cores, collected during RIKEAU 2002 cruise on board r/v Thalia, on the shelf of the Bay of Biscay
-
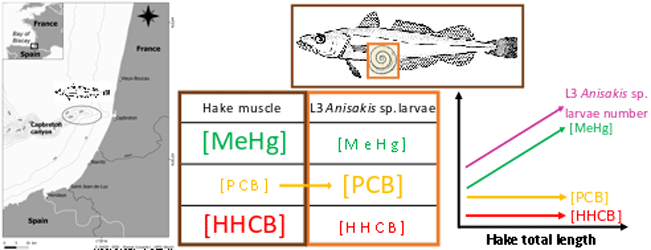
This dataset gathers isotopic ratios (carbon and nitrogen) and concentrations of both priority (mercury species and polychlorinated biphenyls congeners) and emerging (musks and sunscreens) micropollutants measured in a host-parasite couple (hake Merluccius merluccius muscle and in its parasite Anisakis sp) from the south of Bay of Biscay in 2018. In addition, the hake infection degree measured as the number of Anisakis sp. larvae was added for each hake collected.